千家寨野生古茶树叶片解剖结构和化学组分计量特征对海拔梯度的适应(五)
叶肉细胞是寨野征对植物叶片进行光合作用的主要部位,发达的生古栅栏组织既有利于提高植株的光合作用及水分利用率,从而提高植物的茶树抗旱性,还能避免叶肉细胞组织被强光照灼伤(乌佳美等,叶片2019)。不同海拔野生古茶树叶片的解剖结构叶肉组织均分化为明显的栅栏组织和海绵组织,这是和化海拔典型的“异面叶”解剖结构特征,在植物进化过程中异面叶比等面叶更能适应高海拔环境,学组这是分计植物长期适应的结果(赵学鹏等,2019;戴叶辉等,2012)。本研究中,量特野生古茶树叶片栅栏组织厚度、梯度海绵组织厚度及叶肉厚度在其分布的寨野征对中间海拔A2(2200m)拥有最大值,说明在该海拔野生古茶树的生古光合作用最强,更有利于有机物的茶树形成积累,而且该海拔土壤碳源较为丰富,叶片参与有机碳循环过程的解剖结构根际胞外酶和土壤微生物未能及时分解土壤碳也会促使植物积累较多的养分(自海云等,2020)。另外,由于低温和强光照的影响,生长于高海拔的植物,其叶肉细胞普遍表现为“栅栏组织增多变厚、海绵组织减少”(孙会婷等,2016)。而本研究结果显示,叶片海绵组织厚度和栅栏组织厚度均在最高海拔有所增加,而在A3和A4海拔的栅海比亦高于A1和A2海拔,说明高海拔下叶片更倾向于分化更多的栅栏组织,这和前人的观点基本一致。此外,野生古茶树在其分布的最高海拔(2500m)CTR、P/S显著大于最低海拔(2050m)的CTR、P/S,这很有可能是野生古茶树为了提高自身的光能利用率而做出的适应性响应,说明野生古茶树能够很好的适应该海拔较强的光照环境。本研究中,叶片结构疏松度(SR)在海拔2500m拥有最小值,这和钟悦鸣等(2017)对胡杨的研究结果一致,栅海比与叶片结构疏松度同植物的抗寒性有一定的关系,植物叶片栅海比越大、叶片结构疏松度越小,其抗寒抗旱性越强,而且能够保持相对稳定。叶脉主要为植物提供支持、水分传递和光合碳水化合物,其厚度变化直接影响到植物叶片的水分状况和光合产物的运输(Lawrenetal.,2013)。与最低海拔相比,野生古茶树在最高海拔的主脉厚度和叶片主脉突起度均显著增加,说明野生古茶树叶片主脉在最高海拔(2500m)较为发达,这和王元元等(2016)对草血竭的研究结果一致。这主要是由于随着海拔的升高,植物叶片可能出现生理性缺水的现象,而叶脉增大说明植物运输水分及营养物质的能力在增强(范志霞等,2019);叶脉厚度增大,还能提高植物的机械支持能力,防止因海拔升高风力增强而对植物叶片造成机械性损伤。 植物体内C、N、P元素的组成、分配及其周边环境共同决定了植物的生长发育和营养水平,因此环境变化容易导致植物叶片化学组分计量特征具有差异性。本研究中,各海拔野生古茶树叶片C、N、P含量(514.03~540.43、21.06~24.45、2.09~2.81g·kg-1)普遍高于全球陆生植物叶片的C、N、P平均含量(464.00、20.60、1.99gkg-1)和中国陆生植物叶片N、P平均含量(20.20、1.46g·kg-1)(汪宗飞等,2018),表明千家寨野生古茶树叶片有机化合物含量较高,氮磷元素较为丰富。其中,叶片C含量呈现先增后降再增的变化趋势,这和张小芳等(2019)对火绒草的研究结果一致。在2200m处叶片C含量高可能是因为该海拔处水热条件良好,植物光合作用及碳同化能力较强,生长势强并积累含C有机物较多;而植物为了适应逆境会积累较多的非结构性C(如淀粉、可溶性糖)以保护植物体不受损伤(洪江涛等,2013),因此叶片C含量在2500m处有所增加。与叶片C含量不同,野生古茶树叶片N、P含量均随海拔的增加先降低再升高,这和以往研究结果一致(Hikosakaetal.,2010)。N主要以蛋白酶的形式参与植物光合作用,随海拔上升,光照增强,蛋白酶消耗较多,因此N含量逐渐减少;而随着海拔继续升高,温度逐渐降低,较高的N、P含量有利于减缓植物在低温环境下的生理过程(Fanetal.,2016;Zhongetal.,2015)。而叶片P含量的增加,还可能是随着海拔的升高,降水量增多导致土壤水分含量增加,加快了土壤母质矿质风化的速率,使得土壤中的有效P含量增加(刘俊雁等,2020;Zietal.,2020)。 叶片C/N和C/P比值可衡量植物吸收养分的碳同化能力,在一定程度上能反映植物的营养利用率。本研究发现,野生古茶树叶片C/N、C/P在海拔A2显著增至最大水平,说明在该海拔野生古茶树有着较高的氮、磷利用效率,这可能是叶片高C含量和低N、P含量共同作用的结果,说明植物为了更好地适应不同海拔小生境,可通过均衡叶片化学组分的含量及保持其化学计量比值平衡等方式来维持植物体的内稳性(杜满义等,2016)。另外,叶片N/P常用来判断植物养分限制因子,N/P<14,植物生长主要受N限制;N/P>16,植物生长主要受P限制;14 可塑性指数PI和变异系数CV是衡量植物适应能力的两个指标,叶片的可塑性指数和变异系数越大,说明植物对环境的适应能力越强(Ashtonetal.,2011;Valladaresetal.,2000)。本研究发现,叶片上下角质层厚度、下表皮厚度以及主脉厚度的PI值较大,而C含量的PI值较小,说明野生古茶树叶片的上下角质层厚度、下表皮厚度和主脉厚度受到海拔环境变化的影响较大,而海拔对叶片C含量的影响微弱。此外,千家寨不同海拔野生古茶树的叶片解剖结构指标的变异系数在0.1~1.0范围内,说明存在中等变异(Ashtonetal.,2011)。叶片N含量的变异系数(0.07)低于P含量的变异系数(0.12),这和贺合亮等(2017)研究结果一致,植物体内N比P具有更强的内稳态系数,因此N含量在植物中的变异性小于P含量。本研究还发现,野生古茶树叶片各解剖结构的可塑性指数与变异系数普遍较高,说明野生古茶树叶片解剖结构与生态化学计量特征具有相对的海拔变异性和稳定性,野生古茶树叶片解剖结构对环境变化表现出较大的可塑性,间接表明该物种对不同海拔小生境有较高的潜在适应能力。另外,PCA分析结果表明,前两主成分轴解释了总变异的87.66%(图6A),其中叶片解剖结构等相关指标贡献率较大,主要表征千家寨野生古茶树抵御外界不利环境和有效利用水热资源及土壤养分的能力,第一主成分主要突出反映植物抵御外界干扰的能力,第二主成分突出反映植物对光照和水分等异质性环境因子的资源利用策略。千家寨野生古茶树叶片解剖结构和化学组分计量特征在不同海拔环境表现出的差异,也体现了茶种植物对多种环境的适应能力。 本研究从形态结构及生理生态角度对野生古茶树随垂直气候变化的变化进行了探讨,研究结果可为深入理解野生古茶树对环境的适应机制提供科学依据。本研究所选野生古茶树分布范围海拔跨度较小,且仅对其叶片解剖结构和化学组分计量特征方面进行了比较分析,具有一定的局限性,今后可以增加野生古茶树的采样点,对其叶片代谢产物、生理生化乃至分子机制等做更深入的研究,以期为古茶树资源的合理保护、开发和利用提供更全面更科学的指导。 千家寨4个海拔野生古茶树群体叶片解剖结构和化学组分含量及其计量比差异显著,主要受环境变化的影响。野生古茶树叶片养分含量整体偏高,在生长过程中不受N、P元素的限制,其叶片解剖学特征的变异性较强。中间海拔2200m为野生古茶树的适生区,其光合碳同化及生长代谢较为旺盛,而在海拔2500m,野生古茶树具有较强的保水能力和抗旱抗寒能力。 声明:本文所用图片、文字来源《生态学杂志》,版权归原作者所有。如涉及作品内容、版权等问题,请与本网联系删除。 相关链接:细胞,植物,环境3.2野生古茶树叶片化学组分计量特征对海拔的响应

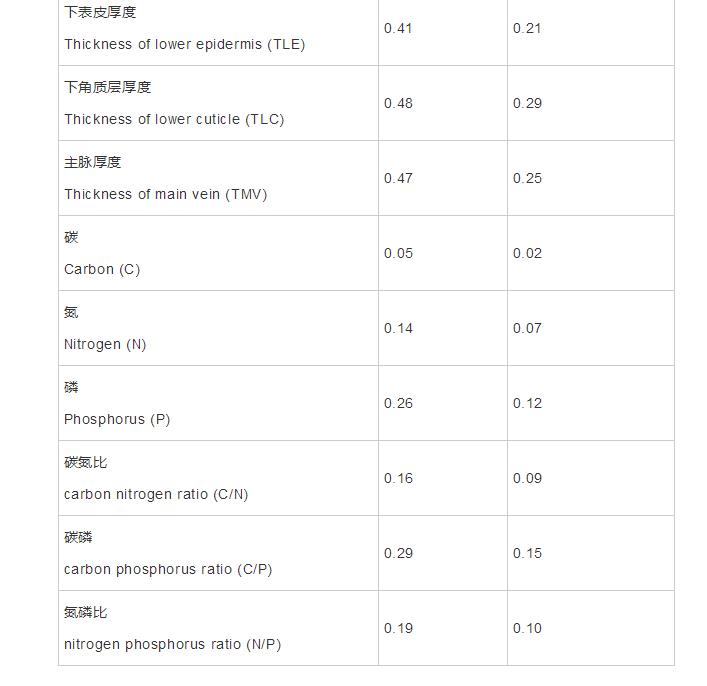
3.3野生古茶树叶片解剖学特性和化学组分计量特征对海拔的适应能力
4结论
- 最近发表
- 随机阅读
-
- 高盐稀态酱油中生物胺的差异性分析(三)
- 稱中國正在欺凌鄰國、威脅台灣,北約首次將「來自中國的挑戰」納入戰略文件
- 前日相安倍晉三街頭演說遭霰彈槍槍擊,當場「心肺停止」急送醫院
- 倪匡逝世享年87歲,沈西城:香港難以再找到這樣的作家
- 张八岭镇未雨绸缪扎实做好防汛备汛工作_
- 前中華籃協副秘書長王啟先逝世享壽72歲,曾推動男籃換血催生曾文鼎、林志傑等「黃金世代」
- 從亞馬遜到Google紛紛投入,一篇看懂「零售媒體聯播網」(RMN)
- 民進黨提名即將進入「深水區」:花東縣長人選將出爐,雙北重中之重難喬定
- 被困K396次列车乘客,徒步9小时求生只想给家里报平安
- 「已經拿到想要的了」,土耳其不再反對,北約正式邀請芬蘭和瑞典加入聯盟
- 從佛法角度定位「天上聖母」:媽祖是觀音菩薩的化身嗎?修行境界有多高?
- 全球缺水危機加劇,海上核動力「除鹽工廠」能幫助解決問題嗎?
- 特种钨合金成分分析标准物质研制(二)
- 日本科學家用冷凍乾燥體細胞成功複製實驗鼠,未來有望保護物種免於滅絕
- 《陳設美好的生活》:陳設的四大原則,以及選購居家陳列物與生活道具的十個關鍵
- 「好看」背後的設計思維:深度分析蘋果發表會簡報排版技巧
- 碳酸钠标准品:精准测量与分析必备
- NBA刺青文化的演變(下):刺青成為個性簽名,球員刺上對自己具有重大影響的人事物
- 稱中國正在欺凌鄰國、威脅台灣,北約首次將「來自中國的挑戰」納入戰略文件
- 從史蒂芬金《岩堡城》,看美國的「恐怖」(上):劇中的恐怖,來自刻意被遺忘的歷史
- 搜索
-
- 友情链接
-