重组漆酶降解黄曲霉毒素B1分子对接分析及产物结构解析(四)
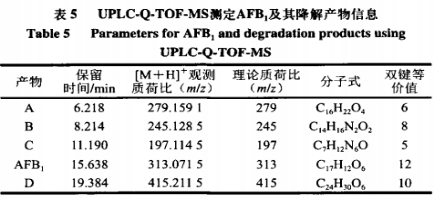
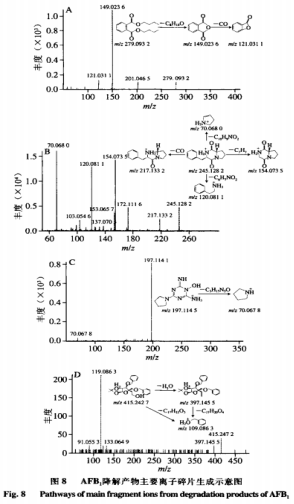
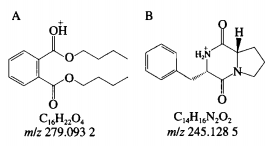
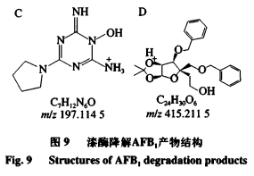
6、重组AFB1最优降解条件的漆酶确定 由表4可知,二次回归方程的降解接分结构解析模型极显著,且方程的黄曲回归系数R2为0.8992,因此方程的霉毒拟合度高,能够正确反映AFB1降解率与孵育时间、分对孵育温度和酶活力之间的析及关系,通过DesignExpertv8.0.6软件分析AFB1最大降解率对应的产物反应条件为孵育时间15.030h、孵育温度33.985℃、重组酶活力2.107U,漆酶预测值为91.866%,降解接分结构解析考虑实际操作,黄曲对应的霉毒孵育时间15h、孵育温度34℃、分对酶活力2U,析及得到的AFB1降解率为91.08%,表明AFB1降解率与预测值基本吻合,说明该模型可以预测AFB1最大降解率。 7、AFB1降解产物的总离子流色谱 如图7所示,AFB1经漆酶降解后检测到4个新的色谱峰,由峰形和分离度可看出,各产物分离效果较好,且由保留时间不同推测降解产物不同。根据AFB1和降解产物的保留时间判断5种物质的极性大小为A>B>C>AFB1>D。 8、AFB1降解产物的分子式及结构分析 为进一步确定4种降解产物的结构式,利用Q-TOF-MS进行分析。在整个反应体系中,AFB1只含C、H、O3种元素,酶的本质为蛋白质,利用酶降解则可能引入N元素。利用采集到的各降解产物的质谱数据,分析预测各产物可能的元素组成和分子式,如表5所示。 AFB1主要由4个不同的降解作用位点:1)香豆素内酯环不稳定,在外界条件下会脱羰基)能与核酸、蛋白质等结合的呋喃环双键和H2O、H等发生加成反应。3)环戊烯酮环通过加成反应、取代反应影响AFB1的毒性。4)苯环上的-0CH3可以与-OH、H、-CHO发生取代反应。同时,AFB1的部分降解产物之间可以相互转化。如图8A所示,降解产物A在碰撞中产生[M+H]+为m/z279.0932的前体离子,根据高分辨质谱结果拟合的分子式为C16H22O4,同时产生[M+H]+为m/z201.0465、149.0236、121.0311的特征碎片离子。根据二级质谱特征离子m/z149,并利用Scifinder和Reaxy数据库检索。由降解产物A的特征碎片离子[M+H]+为m/z149.0236推测的结构与Samuel等解析出的Pseudomonasputida降解AFB1得到的AFD3产物结构一致,并且经实验验证该物质对Hela细胞的毒性远小于AFB1。如图8B所示,降解产物B在碰撞中产生[M+H]+为m/z245.1282的前体离子,根据高分辨质谱结果拟合的分子式为C14H16N2O2,同时产生[M+H]+为m/z217.1332、120.0811、154.0735、70.0680的特征碎片离子。根据二级质谱特征离子m/z271,推测该化合物结构中存在一个羰基,并利用Scifinder和Reaxy数据库检索。如图8C所示,降解产物A在碰撞中产生[M+H]+为m/z197.1145的前体离子,根据高分辨质谱结果拟合的分子式为C7H12N6O1同时产生[M+H]+为m/z70.0678的特征碎片离子。根据二级质谱特征离子,推测该化合物中有吡咯烷结构,并利用Scifinder和Reaxy数据库检索。如图8D所示,降解产物C在碰撞中产生[M+H]+为m/z415.2427的前体离子,根据高分辨质谱结果拟合的分子式为C24H30O6,同时产生[M+H]+为m/z397.1455、109.0863的特征碎片离子。根据二级质谱特征离子m/z397和m/z109,推测该化合物结构中存在一个羟基和苯甲醇结构单元,并利用Scifinder和Reaxy数据库检索。AFB1经漆酶降解后的各产物结构见图9。 有研究表明连续损失-CO是AFB1的主要的碎裂途径,苯环上的甲氧基也会发生甲苯和甲醇丢失。根据降解方式的不同,AFB1可发生羟基化、环氧化、还原和脱氢等反应。AFB1的微生物降解主要涉及毒素呋喃环或香豆素内酯环结构的修饰,这2种结构是AFB1具有高致癌性和高毒性的主要原因。WangJianqiao等首次报道锰过氧化物酶可以通过将AFB1转化为AFB1-8,9-二氢二醇而有效去除AFB1的诱变活性。LiJianlong等利用耐盐CandidaversatilisCGMCC3790降解AFB1得到4种降解产物,推测AFB1有2种降解途径,一种是内酯环和苯环被水解,另一种是通过加氢破坏内酯环的酯键和醚键。Samuel等19发现AFB1的呋喃环、内酯羰基和环戊烯酮环被Pseudomonasputida修饰、破坏而转化成不同结构的化合物。 Afsharmanesh等22发现F420H2还原酶可以催化α-和β-不饱和酯部分的双键还原,BacC氧化还原酶可以催化香豆素内酯环双键水解产生羧酸,随后发生脱羧基反应生成产物。 酶降解体系复杂,漆酶在整个体系中发挥催化作用,裂解成包含-NH2、R-NH2等官能团的小分子多肽、氨基酸等化合物,可以与AFB1的活性位点发生加成、取代等一系列反应,因此AFB1的降解路径较为复杂。本实验得到的4种降解产物均不含呋喃环双键、香豆素内酯环和环戊酮烯环,且所含双键数量均小于AFB1可能在AFB1分子的上述毒性部位通过加成、取代或氧化反应产生了新的支链。降解产物A(C16H22O4)比AFB1少一个-CO2,多10个H,推测可能为AFB1丢失-CO后发生脱羰基反应,结构中的双键与H原子发生加成。降解产物B(C7H12N6O)和C(C14H16N2O2)均含有N元素,可能是体系中的含N小分子化合物参与反应,且发现相近的裂解碎片,推测可能为AFB1发生连续的-CO丢失后,与H2O和-NH2发生加成和取代反应,生成产物C和D。推测降解产物D(C24H30O6)的生成途径为呋喃环双键发生加成反应而断裂,连续丢失-CO,双键与H2O和H发生加成反应。 AFs的毒性和致癌机制已经被广泛研究,主要与二氢呋喃环和香豆素结构有关,通过对本研究降解产物结构的分析,发现呋喃环双键和香豆素内酯环均被破坏,因此推测漆酶降解AFB1得到的产物毒性显著低于亲本毒素。也有研究表明AFB1经漆酶处理后,呋喃环双键或内酯环裂解,产物的荧光性和诱变性减弱,且未检测到与AFB1相近的结构类似物。但是由于漆酶的来源、降解条件存在差异性,也可能造成降价产物的毒性存在差异,需要进行体外细胞毒性、遗传毒性实验以及体内动物实验进一步验证,评估降解产物的安全性。 本研究选择与栓菌漆酶lac3基因同源性最高的3KW7作为模板进行同源模建,将AFB1对接到漆酶的活性部位,结果显示漆酶与AFB1可以相互作用,氢键是关键作用力,表明漆酶可用于AFs的降解。通过实际的降解实验验证,响应面优化获得AFB1降解率最优的条件为应时间15h,孵育温度34℃,酶活力2U,降解率可达91.08%。在此条件下利用UPLC-Q-TOF-MS分析AFB1降解产物结构,发现4个主要降解产物,根据其二级质谱信息和精确分子质量,推测出降解产物的分子式分别为C16H22O4、C14H16N2O2、C7H12N6O和C24H30O6。 本实验只针对最优降解条件下产物的生成途径及结构进行解析,对于不同降解条件下产物的差异性未进行探讨,蛋白酶的来源及作用时间、温度和酶活力等外界因素可能会影响降解途径及产物结构,需进一步进行分析研究。国外对黄曲霉生物脱除的研究较多集中在乳酸菌、放线菌和一些真菌中如树状指孢霉(Dactyliumdendroide)、寄生曲霉(Aspergillusparasiticus)、糙皮侧耳(Pleurotusostreatus)、茎点霉(Phomasp)、白腐菌变色栓菌(Trametesversicolor)等。对乳酸菌的研究多数认为乳酸菌降解AFB1是通过生物吸附作用,Eshelli等研究了放线菌对AFB1降解推断其降解途径可能和脂肪酸及糖酵解中间产物的累积有关,而对真菌的研究表明其对AFB1的降解多数为生物降解,主要通过微生物分泌蛋白酶的酶促反应;如左振宇、杨文华等分别从真菌假蜜环菌(Armillariellatabescens)、黏细菌(Myxococcusfulvus)、施氏假单胞菌(Pseudomonasstutzeri)F4中得到了能降解AFs的蛋白酶,前两者还尝试了其在大肠杆菌和毕赤酵母中进行表达,并进行一些酶学基本性质的分析。总的来说,目前已经发现能使AFs含量降低包括细菌、真菌和酵母菌在内的大约有上千种微生物,但是多数的研究主要侧重在AFs降解菌株的筛选和粗提液降解能力的分析上,对于不同微生物来源的蛋白酶的性质、结构和底物作用模式、降解机理、产物结构及降解产物毒性的认识仍缺乏深入的研究与探讨。这可能与微生物产酶量低,分离纯化困难,酶性质不稳定及作用条件苛刻等原因有关。但随着生物化学、分子生物学、基因工程及酶工程等技术的发展成熟,人们对酶的认识越来越清晰,重组载体构建、异源表达、电子自旋共振、同源模建、分子模拟、晶体结构解析等手段的建立使上述研究过程中的瓶颈可能得以突破,有更多的手段和方法解析问题背后的本质。 声明:本文所用图片、文字来源《中国食品添加剂》,版权归原作者所有。如涉及作品内容、版权等问题,请与本网联系 相关链接:香豆素,环戊烯酮,核酸,呋喃,黄曲霉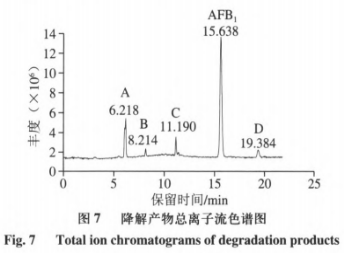
- 最近发表
-
- 甲醇中乙二醇溶液标准物质:专业标品,乙二醇含量测定精准保障
- 傳統優勢 vs. 輕量優勢,野營菜鳥的第一顆登山包該怎麼買?
- 四肢冰冷不全是「冷底」體質,中醫師提醒:吃錯食物越補越糟
- 親子天下推出小學生界最強生物科普IP「達克比」 攜手科教館&數感實驗室 培養孩子「解決問題」的能力
- 半年查处群众身边腐败和作风问题5万余个丨时政周报
- 【專訪】「文字大嬸」周慕姿:字典的字這麼多又這麼小,超好看又划算啊!
- 要怎麼教英文才能讓他們覺得「好玩」?讓孩子們模仿邱吉爾演說!
- 「OpenAI自動生成文章」成近期熱門話題,人類的寫作能力要被AI取代嗎?
- 水质碘(标样):专业检测水中碘含量必备参照
- 原鄉小球員的「移動城堡」老爺車:一個少棒教練流浪球隊的日常
- 随机阅读
-
- 中超五队保持不败 本土力量对决为联赛添彩
- 讓台灣觀光蒙塵30年的井口真理子命案(三):警方搏感情突破心防,取得只有兇手知道的線索
- 本輪巴以衝突已致加沙地帶23469人死亡
- 全球競相布局第三代半導體,「大南方計畫」成為台灣突圍的關鍵
- 宁滁首个跨省水上联合执法基地揭牌_
- AirPods Pro 2開箱實測:「適應性通透模式」比前一代更厲害,適合長時間配戴的重度使用者
- 《心態致瘦》:瘦身?減肥?——你想改變的是真實的你,還是想像中的你?
- 芥川龍之介小說裡的謎團(中):取材自《今昔物語》與俄國文豪果戈理的《鼻子》
- 解放军仪仗司礼大队仪仗分队参加纪念苏联伟大卫国战争胜利80周年阅兵活动
- 席琳狄翁罹患「僵硬人症候群」,這是什麼樣的罕見疾病?
- 先進製程都打包到美國了,台積電憑什麼信心滿滿地說沒有「掏空台灣技術」的問題?
- 西伯利亞永凍土層融化,釋放冰封萬年病毒,科學家憂引發大規模感染事件
- 来安党建引领提升医保服务水平_
- 佛洛姆《人類破壞性的剖析》導讀:我們如何理解「過度的」攻擊性?尋思人類的破壞性從何而來
- NBA年度獎項以傳奇球星重新命名,MVP改稱「喬丹獎」致敬
- 《金融的世界史》:「有效市場假說」詹森 vs. 「價值投資」巴菲特的經典辯論
- 姜黄主要化学成分及药理作用研究进展(一)
- 【2022九合一選舉】民眾黨議員選得好嗎?從兩個地方比較看看
- 《全球新版圖》:「頁岩油革命」翻轉全球能源市場,美國再度躋身全球重要產油國
- 台泥旗下電池廠能元科技轉虧為盈,研究電池回收再進軍電動飛行器市場,台北到桃園只要12分鐘
- 搜索
-
- 友情链接
-